

![]()
CARATTERISTICHE
METAMORFOSI
La metamorfosi è un processo di sviluppo per cui l'individuo assume il suo aspetto definitivo dopo una o più fasi in cui le sue sembianze sono completamente diverse.



La metamorfosi degli Anfibi è un processo mediante il quale un girino si sviluppa in un organismo adulto, nella maggior parte dei casi terrestre. Questo comporta profonde modificazioni morfologiche esterne accompagnate da cambiamenti fisiologici interni.
La trasformazione della larva in anfibio adulto, la metamorfosi, è legata ad un cambiamento ambientale e/o a profonde modificazioni operatisi nel sistema organico e nella struttura corporea.
I mutamenti esterni possono esser seguiti chiaramente nelle larve di Anuri: inizialmente si allungano nettamente gli arti posteriori, poi compaiono quelli anteriori, che si sono sviluppati nella camera branchiale e che fino a quel momento rimanevano nascosti dalla piega opercolare; la coda si riduce fino a scomparire completamente , mentre la pelle si adatta alle esigenze poste dalla vita terrestre e forma ghiandole mucipare e velenifere pluricellulari, assumendo al contempo la colorazione che caratterizza gli individui adulti. Becco e dentini cornei scompaiono, per cui la bocca del girino si trasforma in quella tipica dell'adulto; gli occhi si fanno più grandi e vengono protetti da palpebre adattandosi alla visione aerea, mentre anche gli altri organi di senso subiscono analoghe modificazioni: nell'apparato uditivo, ad esempio, si sviluppa la membrana timpanica che chiude dall'esterno l'orecchio medio.
A queste trasformazioni se ne aggiungono altre non immediatamente visibili: le branchie scompaiono ed entrano in attività i polmoni; sistema circolatorio, muscolatura e scheletro si modificano; il tubo intestinale, che nel girino aveva la lunghezza nove volte superiore a quella del corpo, s riduce fino a raggiungere la stessa dimensione del corpo, adattandosi contemporaneamente all'alimentazione zoofaga; i reni subiscono modificazioni a conclusione delle quali non secernano più ammoniaca, bensì urina.
La metamorfosi provoca anche negli Urodeli cambiamenti altrettanto profondi, seppur meno vistosi, in quanto l'aspetto esteriore rimane sostanzialmente inalterato.
Lo stadio larvale presenta durata diversa da specie a specie e può essere ridotto o prolungato dall'influsso di temperature più o meno elevate e da altri fattori ecologici: spesso copre l'arco di settimane o mesi, oppure di un anno. Alcune specie possono iniziare la metamorfosi già 12 giorni dopo la deposizione delle uova, mentre altre iniziano a trasformarsi in adulti solo dopo 14 o 26 mesi; alcune specie poi, rimangono allo stadio larvale anche per anni.
DALL'UOVO ALL'ADULTO.........
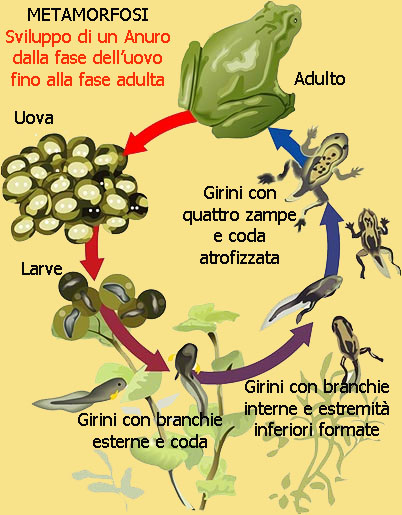
Poco dopo la fecondazione, il nucleo dell'uovo si segmenta in due nuclei più piccoli che si separano; le divisioni dei nuclei "figli" e relativi protoplasmi si susseguiranno da questo momento in ritmo incalzante fino a quando l'uovo primitivo non sarà una palla cava rivestita di cellule. Durante questi fasi le dimensioni dell'uovo non mutano di molto ciò che cambia è il numero di cellule, ciascuna con il suo nucleo e protoplasma. Tutto ciò avviene nei primi due o tre giorni, dopo la sfera si allunga e in essa si distinguono una testa ed una coda mentre le altre cellule dell'embrione si sono organizzate a formare i primi organi della larva. Dopo 10 giorni, la gelatina che ricopre l'embrione si scioglie permettendo al nuovo essere di uscire nell'acqua. A questo punto il processo si differenzia tra uova di urodeli e uova di anuri.
Le larve di urodeli sono sottili, snelle e presentano alla schiusa gli abbozzi branchiali, occhi e due tubicini che servono come organi di attacco. Subito le branchie si completano, sul tronco compare una cresta dorsale,la coda si orna di membrane natatorie. Si formeranno prima gli arti anteriori poi i posteriori. la bocca munita di dentini fornirà alla larva le prime prede animali, essendo già fin da ora carnivori. Le restanti trasformazioni a carico dei vari organi, porteranno la larva acquatica alla vita terrestre propria dell'adulto.

I girini degli Anuri invece, appena usciti dall'uovo si presentano inattivi, privi di bocca e di occhi per i primi giorni, presentano solo un organo adesivo ventrale che gli permette di fissarsi alle piante sommerse.

Dopo otre giorni si apre la bocca, le branchie esterne iniziano a funzionare ricevendo sangue e consentendo la respirazione, la coda si competa di due membrane che ne aumentano le capacità motorie.

Dopo circa tre settimane le branchie esterne lasciano il posto a quelle interne, la dimensione del corpo aumenta e il girino è in grado di nuotare e alimentarsi; il girino è erbivoro e il becco e i dentini servono per raschiare la vegetazione acquatica.

A otto settimane le zampe posteriori si completano e la coda inizia a regredire. Spesso il girino sale in superficie per assumere aria nei piccoli polmoni che stanno formandosi. Dopo un paio di settimane comparirà la prima zampa anteriore e la coda sarò ormai riassorbita. La piccola rana è pronta a lasciare lo stagno per respirare ossigeno con i suoi polmoni. La maturità sessuale sarà raggiunta in un secondo tempo; la rana comune è in grado di generare dopo 4 anni dalla metamorfosi, altre specie possono essere più precoci o più tardive in relazione anche alle condizioni climatiche e ambientali.
NEOTENIA

La neotenia è il mantenimento di alcuni aspetti della fase larvale durante la fase adulta di un animale.
Negli anfibi, la persistenza dei caratteri larvali è
accompagnata al mantenimento degli animali nell'ambiente acquatico e all'aumento
della loro temperatura corporale. Il passaggio dello stadio larvale a quello
adulto è contraddistinto dalla metamorfosi attraverso la quale, nel caso degli
anuri, acquisiscono la capacità di respirare e muoversi nell'ambiente terrestre
convertendosi in rane adulte con capacità riproduttive.
La neotenia si sviluppa maggiormente tra gli urodeli,
il quale ciclo vitale non è chiaro come quello degli anuri essendo le larve meno
distinte rispetto agli adulti.
Si distinguono diversi tipi di neotenia:
Neotenia accidentale o geográfica:
Neotenia nella quale, dentro una medesima specie, la maggior parte degli animali realizzano una metamorfosi completa e le forme che si mantengono in stato larvale sono rare e geograficamente localizzate.

Ciò avviene nel caso della Salamandra Tigre del Nord America (Ambystoma tigrinum) e nell'Ambystoma gracile distribuito a largo delle coste del Canada, in diversi tritoni del Sud Europa (Triturus helveticus) e negli Stati Uniti nel Notophthalmus viridescens. Riguardo l'Ambystoma gracilis si ha comprovato che alcune colonie soffrono di neotenia e ciò può essere legato a quei fattori esterni che influiscono sopra l'apparizione della neotenia in una specie inizialmente non geneticamente legata a questo fenomeno.
Neotenia facoltativa:
Presente in specie nelle quali alcun individui si metamorfizzano completamente in condizioni naturali, mentre altri, vivendo nel medesimo ambiente, persistono e si riproducono nello stadio larvale. Questo tipo di neotenia è influenzabile, e ciò significa che, mediante differenti tecniche si può conseguire che un individuo neotenico si metamorfizzi e raggiunga lo stadio adulto. Il miglior esempio di questo tipo si ha con l'Ambystoma mexicanum, Axolotl, endemico del Messico.

Si sono conclusi distinti esperimenti nei quali si è preso un gruppo di individui e nelle stesse condizioni
alcuni subiscono metamorfosi spontanea e altri permangono in stadio larvale per
tutta la loro vita. Anche i discendenti degli animali metamorfizzati non possono
concludere la metamorfosi e permangono in stato neotenico malgrado i loro
progenitori fossero salamandre adulte di Ambystoma mexicanum.
Confrontando un esemplare di A. mexicanum metamorfizzato con uno in
stadio neotenico possiamo riscontrare importanti differenze morfologiche.
Neotenia obbligatoria:

Certe specie non subiscono mai una metamorfosi completa e permangono sempre in
uno stato larvale più o meno profondo. Esempio di ciò il Necturus
maculosus, il Proteus anguinus, l'Andrias japonicus, l'Amphiuma
means e lo Pseudobranchus sp.
Caratteristiche morfologiche - anatomiche - istologiche degli anfibi neotenici
La neotenia si può mantenere in parte
o nella sua totalità negli individui neotenici in modo da avere diversi gradi ed
è possibile osservare ciò nelle specie già nominate.
Una metamorfosi quasi completa si realizza nell'Andrias japonicus, una rete densa di capillari sanguigni con un "foglio" respiratorio si installa intorno al derma e sostituisce, alla volta dei polmoni, l'assenza di branchie in questa specie.
Negli Amphiumas, la metamorfosi è tuttavia importante ed elimina numerosi
caratteri larvali. Le branchie esterne spariscono così come i bordi della coda
che aiutano alla propulsione natatoria. Le trasformazioni riguardano la pelle,
la cloaca e la bocca e persistono un paio di fenditure branchiali aperte come le
branchie larvali.

Nei Sirenidi e Axolots, l'organizzazione larvale è appena
modificata e i segni della metamorfosi sono discreti, conservano le tre paia di
branchie esterne e il bordo caudale che li facilita nei movimenti nell'acqua,
però perdono alcune cellule epidermiche.
In alcune specie i caratteri larvali si conservano nella loro totalità e non si
produce alcun passaggio anatomo-morfologico relazionato con la metamorfosi.
Nei Proteus e Necturus, il bordo caudale natatorio persiste così come spesso le tre paia di branchie esterne. Tutte le fenditure branchiali sono aperte e il cranio è poco ossificato. Gli archi branchiali così come la pelle sono uguali allo stadio larvale. I polmoni, che appaiono prima della metamorfosi in tutti gli anfibi, sono sacchi poco sviluppati con funzione idrostatica e la loro funzione respiratoria è molto ridotta.
Nel Triturus helveticus gli individui
soffrono di distinti gradi di metamorfosi così si incontrano esemplari con
neotenia parziale e altri con neotenia quasi totale.
Determinazione ormonale della neotenia
La neotenia negli urodeli è associata ad un attività insufficiente del centro ipotalamo-ipofisotiroideo; ad un funzionamento non corretto quindi dell'ipotalamo, ipofisi e ghiandola tiroidea.
Negli urodeli neotenici si produce un
ipofunzionamento della ghiandola tiroidea che produce quantità minori di ormoni
rispetto alla norma. Tali concentrazioni ormonali sono insufficienti per
produrre le trasformazioni morfologiche e anatomiche in un Axolot, per esempio,
però sono sufficienti per produrre scambi chimici nelle emoglobine e il
passaggio dall'escrezione di ammoniaca a quella dell'urina. In alcuni esemplari
si produce un aumento spontaneo dei livelli di tirossina producendosi così una
metamorfosi completa.
Nel caso degli urodeli neotenici
"obbligatori", si sono studiati gli ormoni e si è visto che sono prodotti ma non
raggiungono il flusso sanguigno. Inoltre gli organi di questi individui non
hanno recettori per questi ormoni e per tanto mai si produrrà un caso di
metamorfosi spontanea ne indotta artificialmente.
Da chiarire il perchè del basso livello di tirossina nel sangue e quanto sia la tiroide colpevole del mal funzionamento.
Si è dimostrato che inducendo la ghiandola, funziona correttamente e si produce la metamorfosi negli individui studiati.
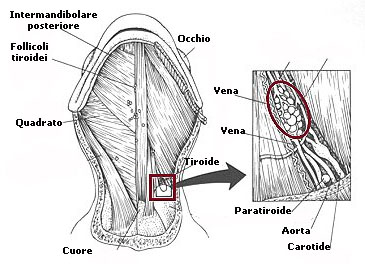
Il funzionamento della ghiandola tiroidea è controllato da un'altra ghiandola, l'ipofisi che si compone di due parti:
la neuroipofisi che proviene dal tessuto nervoso e
la adenoipofisi che proviene dal tessuto della bocca non nervoso. Le due parti
comunicano fra loro, la neuroipofisi regola la adenoipofisi segnalandole il
momento in cui deve secernere gli ormoni.
Uno degli ormoni che secerne l'adenoipofisi è l'ormone tiroideo (TSH), ormone
che va ad influenzare la tiroide e la farà lavorare. La tiroide secerne
tirossina T3 e T4, ormoni ricchi di iodio che presentano una parte fondamentale
nella metamorfosi degli anfibi.
La scarsa produzione di tirossina negli urodeli
neotenici si deve alla
scarsa produzione di TSH da parte dell'adenoipofisi. Le cellule che producono la
secrezione di TSH sono più scarse che negli animali in metamorfosi. In individui
neotenici la TSH non si produce in quantità sufficiente per attivare la
secrezione di tirossina nella ghiandola tiroidea.
Esistono però altri ormoni che influiscono nella metamorfosi, come la prolattina. Si è dimostrato che questo ormone se si presenta in grandi quantità nel plasma sanguigno può produrre neotenia. Produce una diminuzione nella produzione di TSH nell'adenoipofisi producendo a sua volta un calo nella produzione di tirossina.
Risposta dei tessuti agli ormoni tiroidei
Il trattamento di urodeli neotenici con
ormoni tiroidei, induttori della metamorfosi negli anfibi, ha dimostrato che i
tessuti e organi hanno reazioni differenti secondo le specie.
Qualunque sia la dose di ormoni, i tessuti del Necturus e del Proteus risultano incapaci di subire metamorfosi; quindi risulta obbligatori ala neotenia. A quanto pare i loro organi hanno perso i recettori per questi ormoni.
In altri neotenici
obbligatori appartenenti ai Sirenidi, la maggior parte dei tessuti sono
incapaci di metamorfizzare; solo le branchie e la coda mostrano una capacità
limitata e subiscono una parziale atrofia. Tali organi sono poco sensibili agli
ormoni e ne necessitano grandi concentrazioni per attuare una reazione.
In individui
neotenici "facoltativi", come gli Axolotl, tutti i tessuti sono idonei
per gli ormoni tiroidei essendo particolarmente sensibili ad essi. Si produce
una metamorfosi completa con regressione delle branchie e della coda con
differenziazione della lingua e altre modifiche.
Tutto ciò in concentrazioni di tirossina minori a 2x10-6 g/ml.
La sensibilità agli ormoni va diminuendo con l'età; le larve immature rispondono a concentrazioni minori rispetto a quelle mature che necessitano di concentrazioni doppie o triple di ormoni.
Nelle forme neoteniche accidentali come la Salamandra Tigre, tutti i tessuti reagiscono sensibilmente agli ormoni tiroidei. La metamorfosi raggiunge facilmente il suo termine con concentrazioni minori rispetto a quelle utilizzate per esempio per gli Axolotl.

Oltre ai processi
anatomici, spesso avvengono processi di cambi biochimici. La induzione delle
trasformazioni biochimiche attraverso gli ormoni tiroidei sono state studiate
particolarmente negli Axolotl. Sono trasformazioni generalmente molto più
sensibili agli ormoni rispetto le modificazioni morfologiche o anatomiche. In
tal modo con l'iniezione di tirossina si producono nuove emoglobine proprie
dello stadio adulto che favoriscono spesso il passaggio dalla secrezione di
ammoniaca a quella di urina.
Basi genetiche della
neotenia
Bisogna tener conto che la neotenia obbligatoria
e facoltativa degli urodeli è nel loro genotipo, nel loro comportamento
ereditario. Nei neotenici obbligatori, come il Necturus, la scarsa
attività delle ghiandole dell'ipofisi e tiroide sono legate al genotipo.
La neotenia è un gene recessivo
(n), mentre chi realizza la metamorfosi è un gene
dominante in tutti gli individui (M).
Si sono fatti esperimenti incrociati negli Ambystoma mexicanum e Ambystoma tigrinum, incrociando un A.mexicanum neotenico con un A.tigrinum:
| Incrocio di individui Omozigoti | Risultati | ||||
|
Generazione
Parentale |
|||||
| A.mexicanum | X | A.tigrinum | |||
| nn | MM | ||||
| Generazione F1 | |||||
| 100% | Mn | 100% | Metamorfizzati | ||
| Incrocio di individui Eterozigoti | |||||
|
Generazione F1 |
|||||
| Mn | X | Mn | |||
|
Generazione F2 |
75% | Metamorfizzati | |||
| 50% Mn | 25% MM | 25% nn | 25% | Neotenici | |
Possiamo osservare la tabella dell'esperimento.
Seguendo la Legge di Mendel, si incrociano individui omozigoti (due geni uguali per il medesimo carattere), uno omozigote recessivo (neotenico, A.mexicanum) e l'altro omozigote dominante (metamorfizzato, A.tigrinum).
Il risultato è tutta una generazione di individui metamorfizzati (100%).
Il loro genoma è Mn, però, come il gene dominante è quello della metamorfosi, ed è precisamente quello espresso nell'animale, l'altro, rimane custodito dentro il loro genoma.
Incrociando gli individui di questa prima generazione (F1) tra loro incroceremo individui Mn e il risultato è che il 75% si metamorfizzano, mentre il 25% presentano neotenia.
Perchè un individuo sia neotenico debbono unirsi in un medesimo animale, i due geni della neotenia, perciò lo si considera come un gene recessivo. Solo un 25% della seconda generazione ( Generazione F2 ) sarà neotenico.
Fattori esterni
Importante però osservare anche altri fattori esterni in relazione alla neotenia, se un individuo geneticamente non neotenico viene posto a squilibri ambientali allo sviluppo della sua metamorfosi, è possibile che non la sviluppi. Quindi, il componente genetico è sicuramente una parte fondamentale per lo sviluppo della neotenia ma spesso bisogna tenere in conto i fattori ambientali esterni.
Tre normalmente i fattori
principali:
Temperatura: Il freddo è uno dei fattori più importanti. Riduce le reazioni metaboliche degli individui soprattutto la generazione di ormoni tiroidei e la loro secrezione ai vasi sanguigni. Ciò si verifica soprattutto in zone di ala montagna, torrenti e fiumi di grandi altitudini. Nella neotenia facoltativa,che si ha nell'Ambystoma mexicanum, il maggior componente è il freddo dato che frena il metabolismo tiroideo e diminuisce la sensibilità dei tessuti verso gli ormoni che circolano nel sangue
Assenza di luce: sicuramente influente per quegli esemplari che vivono in grotte come il Necturus e il Proteus
Scarsa mineralizzazione dell'acqua: una scarsità negli ioni come il calcio, sodio e magnesio aiutano a mantenere la neotenia. Una bassa concentrazione di iodio può bloccare il metabolismo della ghiandola tiroidea e arrestare la produzione di tirossina.
Si può sostenere quindi che negli
individui che vivono a grandi altitudini, nei ruscelli di alta montagna, in
grotte e acque salmastre, i casi di neotenia possono essere conseguenza tanto di
fattori esterni quanto di fattori genetici. Al contrario, in animali che vivono
in zone temperate e con fattori esterni ininfluenti per il metabolismo, si può
sostenere che la loro situazione dipende esclusivamente da fattori genetici.